Transport wody - jak to robią rośliny?
Poniższy artykuł został opublikowany pierwotnie w czasopiśmie dla nauczycieli Biologia w Szkole (5/2017):

Tlenek diwodoru lub oksydan to jeden z bardziej powszechnie występujących na naszej planecie związków chemicznych. Oczywiście częściej nazywamy tę substancję wodą, a i jej wzór chemiczny H2O jest rozpoznawalny nawet poza kręgami chemicznymi.
Okazuje się też, że woda jest jedną z najpospolitszych substancji jeśli chodzi o występowanie we Wszechświecie [1]. W Układzie Słonecznym można ją spotkać także poza Ziemią. Według badań stanowi (w postaci lodu) ważny element budowy planety karłowatej Ceres i wielu księżyców planet-olbrzymów, w mniejszych ilościach występuje także na powierzchni lub w atmosferach planet typu ziemskiego [2].
Ziemia jest prawdopodobnie jedynym miejscem w Układzie Słonecznym, gdzie w naturalnych warunkach woda może istnieć w trzech stanach skupienia. Wiemy, że woda w stanie ciekłym pokrywa ponad 70% powierzchni naszej planety. Duże ilości wody są zdeponowane w postaci lodu w pobliżu biegunów geograficznych, a także w lodowcach wysokogórskich. Oczywiście, woda znajduje się także w atmosferze - szczególnie w jej dolnych warstwach. Wiele naturalnych związków chemicznych występuje w postaci tzw. hydratów - zawierają one cząsteczki wody w swojej budowie. Uwięziona w ten sposób woda nazywana jest wodą krystalizacyjną. Zawartość wody włączonej w strukturę minerałów obecnych w płaszczu Ziemi może wielokrotnie przekraczać łączną zawartość wody w oceanach [3].
Ze względu na swoje właściwości chemiczne i fizyczne woda ma wręcz fundamentalne znaczenie biologiczne. Substancja ta zapewnia płynne środowisko niezbędne do prowadzenia wszelkich procesów życiowych - jest np. powszechnym rozpuszczalnikiem czynnych biologicznie związków chemicznych. Woda występuje w przebiegu większości reakcji metabolicznych. Stanowi też środek transportu wewnątrzustrojowego substancji odżywczych, enzymów i innych. Dzięki dużej wartości ciepła właściwego (4189,9 J/Kg∙K) umożliwia wydajną termoregulację. Nie sposób tu wymienić oczywiście wszystkich procesów biologicznych, w które zaangażowana jest omawiana substancja, ale warto nadmienić, że według wielu hipotez to właśnie w środowisku wodnym po raz pierwszy powstało życie.
Ponieważ woda w stanie płynnym - z tego co wiemy - jest konieczna do aktywnego prowadzenia procesów składających się na zjawisko życia, to logiczne jest, że każdy organizm musi posiadać odpowiednie mechanizmy pozwalające mu na pobieranie tej substancji ze środowiska. O ile w przypadku organizmów jednokomórkowych nietrudno wyobrazić sobie te procesy, to u bardziej złożonych stworzeń (które mogą osiągać przecież duże rozmiary) musiały się rozwinąć specjalizowane sposoby pobierania wody.
Pozostawiając na boku organizmy zwierzęce i grzyby, zastanówmy się nad tym, w jaki sposób rośliny mogą pobierać wodę z gleby, a następnie rozprowadzać ją po całym swoim organizmie. Ponieważ woda jest potrzebna wszystkim tkankom, pytanie to jest tym bardziej zasadne w odniesieniu do drzew. Za najwyższe obecnie żyjące rośliny uważa się mamutowca olbrzymiego Sequoiadendron giganteum i sekwoję wiecznie zieloną Sequoia sempervirens - dorastają one do wysokości ponad stu metrów. Wyniesienie wody na takie wysokości wymaga zużycia dużych ilości energii. Jak rozwiązały ten problem rośliny?
Jednym z roślinnych mechanizmów pobierania wody jest parcie korzeniowe. Polega ono na wypieraniu wody przez korzenie do łodyg i liści. Przyczynia się ono do krążenia soków roślinnych, zwłaszcza przed pojawieniem się liści.
Powstaje ono dzięki zwiększeniu stężenia jonów i innych substancji małocząsteczkowych w tkankach rośliny w stosunku do wody występującej wokół korzeni. Jony te muszą być transportowane przez roślinę w sposób aktywny, więc cały proces wymaga zużycia zmagazynowanej energii. Woda wnika do rośliny zgodnie z powstałym gradientem stężeń na drodze osmozy [4].
Wytworzone w ten sposób ciśnienie pozwala na podniesienie słupa wody na dosyć dużą wysokość i dostarczenie jej wraz z rozpuszczonymi w niej solami mineralnymi do górnych rejonów rośliny.
Poza parciem korzeniowym rośliny wykształciły także inny, niewymagający od nich nakładów energii sposób na rozprowadzanie wody po ich organizmach. Aby zbadać ten temat, przeprowadzimy odpowiednie doświadczenia.
Konstrukcja modelu
Po pierwsze, trzeba zbudować odpowiedni model doświadczalny. Musi on pozwalać na zaobserwowanie interesujących nas zjawisk, a także być na tyle uproszczony by jego konstrukcja nie nastręczała trudności w warunkach szkolnych czy domowych. Bardzo ważnym warunkiem jest też oczywiście przejrzystość modelu, czyli łatwość uzyskania potrzebnych danych.
Proponowany przeze mnie model składa się jedynie z dosyć łatwo dostępnych elementów – odcinków cienkiej, szklanej rurki i sznurka lub grubej nici z włókien naturalnych (Fot.1).

Schemat budowy modelu został przedstawiony na Rys.1.
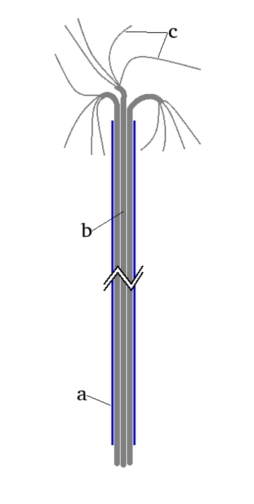
Jak widać, budowa urządzenia jest bardzo prosta. Składa się on z dosyć długiej (przynajmniej 10-15cm) rurki szklanej o niewielkiej średnicy. W wykonanym przeze mnie modelu długość rurki wynosiła 20cm, a jej średnica wewnętrzna ok. 2mm. W ostateczności rurkę szklaną można zastąpić wykonaną z przezroczystego tworzywa sztucznego. Przez rurkę należy przeciągnąć sznurek, a najlepiej kilka splecionych ze sobą sznurków z włókna naturalnego (bawełnianego, lnianego, itp.) tak by dosyć ciasno wypełniły jej światło. Na dolnym końcu rurki sznurki powinny wystawać na niewielką długość, np. 1-2cm (Fot.2).

Na górnym końcu należy natomiast pozostawić kilkukrotnie dłuższy odcinek sznurków, które dodatkowo trzeba rozpleść na pojedyncze włókna, dla zwiększenia powierzchni parowania (Fot.3).

Cały model ma postać widoczną na Fot.4.

Takich modeli w celach porównawczych należy zbudować dwa.
Doświadczenie i obserwacje
Na początku oba modele należy ustawić pionowo w naczyniu z wodą destylowaną, tak by ich dolnej końce wraz z wylotem rurki szklanej były zanurzone (Fot.5). Następnie trzeba nasycić wodą sznurek, nakraplając wodę np. za pomocą pipety pasteurowskiej na górne końce rurek, tak by wewnątrz nich sznurek także został zwilżony na całej długości. Rozplecione górne końce także powinny zostać dokładnie zwilżone.

Górna część jednej rurki wraz z wilgotnymi rozplecionymi końcami powinna zostać szczelnie osłonięta folią z tworzywa sztucznego, tak by ograniczyć parowanie wody z włókien. Drugą rurkę pozostawiamy nieosłoniętą. Gotowy układ doświadczalny przedstawia Fot.6.

Następnie do wody należy dodać barwnik, np. spożywczy lub atrament. Ja zastosowałem wydajny, niebieski barwnik – błękit metylenowy C16H18ClN3S.
Układ doświadczalny od tego momentu powinien się znajdować w ciepłym, dosyć suchym i przewiewnym miejscu. Obserwacje trzeba prowadzić w czasie następnych kilku godzin.
Już po niedługim czasie, czasem nawet po godzinie, można zaobserwować ciekawe zjawisko – poziom wody z barwnikiem wyraźnie się podnosi w rurce. Dzieje się tak jednak tylko w przypadku rurki, której górny koniec nie jest osłonięty. Po kilku godzinach efekt jest bardzo wyraźny – barwnik jest już widoczny na całej długości rurki nieosłoniętej, podczas gdy w przypadku drugiego modelu żaden wzrost poziomu barwnika nie może być zauważony (Fot.7).

Wyniki doświadczenia jeszcze łatwiej prześledzić po wyciągnięciu obu modeli z roztworu barwnika. Można wtedy zauważyć, że w przypadku rurki osłoniętej barwnik dotarł jedynie do miejsca w rurce odpowiadającego poziomowi cieczy w naczyniu, podczas gdy w rurce nieosłoniętej przebył wraz z wodą całą jej długość (Fot.8). Po pewnym czasie rozplecione końce także zostałyby zabarwione.

Zaobserwowane efekty uprawniają nas do stwierdzenia, że w jednym z modeli woda wyparowująca z rozplecionych fragmentów sznurka była zastępowana przez roztwór barwnika wędrujący w górę rurki. Wymagało to wykonania pracy przeciwko sile grawitacji. Praca wymaga jednak wydatku energii. Co więc napędzało obserwowany proces?
Wyjaśnienie
Przypomnijmy sobie na czym polegają zjawiska kapilarne występujące wyraźnie w naczyniach o małej średnicy, np. cienkich rurkach nazywanych też kapilarami (Fot.9).

Wspomniane zjawiska objawiają się m.in. samoczynnym podnoszeniem się poziomu cieczy w kapilarze (Fot.10).

Zjawiska kapilarne nie występują oczywiście jedynie w cienkich rurkach, ale także we wszelkich porowatych materiałach, jak i nasiąkliwych włóknach naturalnych i sztucznych.
Fenomen ten mogliśmy obserwować w zbudowanym przez nas modelu rośliny. W tym przypadku dzięki zjawiskom kapilarnym zachodzącym na cienkich włóknach naturalnych umieszczonych w rurce woda mogła podsiąkać na wysokość większą niż wskazywałby to poziom wody w naczyniu. Wysokość ta nie była jednak na tyle duża, by ciecz mogła pokonać całą drogę wzwyż. Z tego właśnie powodu musieliśmy wstępnie nawilżyć całe włókna.
Włókna w górnej części rurki są odkryte, więc woda jest z nich stale usuwana poprzez parowanie (w przypadku rurki nieosłoniętej). W takim przypadku woda, ewentualnie wraz z rozpuszczonymi w niej substancjami, będzie musiała przepływać z pomocą efektu kapilarnego w kierunku lokalnego spadku zawartości wody z rejonów bardziej nawilżonych. Tak więc obserwowany przepływ wody przeciwko sile grawitacji jest napędzany przez parowanie cieczy z górnych, wolnych końców włókien.
Zauważmy, że na długości całego modelu istnieje pewien gradient, który można zdefiniować poprzez pojęcie tzw. potencjału wody Ψw . Jest to molarna entalpia swobodna cząsteczek wody w układzie. Wartość potencjału wody wyraża się w J/m3 lub Pa. Dla wody niezawierającej domieszek pod ciśnieniem 0,1 MPa oraz w komórkach zachowujących turgor potencjał ten wynosi zero (Ψw =0).
Podniesienie ciśnienia zwiększa wartość potencjału, zaś obecność domieszek obniża go [5]. Przepływ zawsze następuje w kierunku od potencjału wyższego do niższego, co mogliśmy zaobserwować w doświadczeniu.
Taką interpretację potwierdza też fakt, że w przypadku zasłonięcia folią z materiału nieprzepuszczającego wody, a tym samym uniemożliwienia parowania nie zauważyliśmy wyraźnego przepływu cieczy.
Jak się już Szanowny Czytelnik pewnie domyśla, mechanizmy stosowane przez rośliny opierają się na podobnej zasadzie.
Myślę, że należy w tym miejscu przypomnieć, że drewno inaczej nazywane ksylemem jest złożoną tkanką występującą u roślin naczyniowych (Fot.11). Jej główną funkcją jest rozprowadzanie po całej roślinie pobieranej przez korzeń wody i rozpuszczonych w niej soli mineralnych. Duża część komórek wchodzących w skład drewna ma zdrewniałe ścianki, przez co drewno pełni również funkcję mechaniczną [6]. U roślin nagonasiennych w skład drewna wchodzą cewki, zaś u okrytonasiennych naczynia.

Po pobraniu przez komórki włośnikowe korzenia woda przenika w poprzek korzenia do walca osiowego. Transport na tym odcinku odbywa się głównie poprzez apoplast, tj. w porowatej strukturze ścian komórkowych oraz przez przestwory międzykomórkowe. Woda przemieszcza się też drogą symplastyczną - przez protoplasty komórek i plazmodesmy. Z rejonu walca osiowego woda jest transportowana dalej poprzez elementy drewna.
Rozprowadzana w roślinie woda trafia w końcu do liści, poprzez które jest odparowywana, głównie poprzez aparaty szparkowe (roślina ma więc zdolność regulacji intensywności parowania). Proces ten nazywa się transpiracją. Dzięki niej w tkankach liści dochodzi do obniżenia ciśnienia, a więc i potencjału wody w stosunku do pozostałych części organizmu rośliny. Umożliwia to transport wody w naczyniach przeciwko sile grawitacji.
Wartości potencjałów wody kształtują się następująco:
- gleba, Ψw = -0,1 do -0,5MPa
- roślina, Ψw = -0,6 do -1,5MPa
- atmosfera, Ψw = -90MPa [7]
Jak widzimy, transport wody z gleby do atmosfery poprzez roślinę rzeczywiście odbywa się zgodnie z gradientem jej potencjału.
Transport wody w roślinie polega na współdziałaniu parcia korzeniowego i siły ssącej liści powstałej dzięki transpiracji. Wiemy już, że pierwszy proces wymaga od rośliny nakładów energii i jest procesem aktywnym. Drugi zaś można nazwać biernym, ponieważ parowanie odbywa się bez wydatku energii organizmu. W pewnym sytuacjach parcie korzeniowe może mieć jednak znaczenie decydujące – dzieje się tak na przykład na wiosnę, kiedy rośliny nie posiadają jeszcze wykształconych liści, lub przy dużej wilgotności powietrza, kiedy parowanie jest ograniczone.
Literatura:
- [1] Kwok S., Physics And Chemistry of the Interstellar Medium, University Science Books, 2007 powrót
- [2] Dyches P., Chou F., The Solar System and Beyond is Awash in Water, NASA / Jet Propulsion Laboratory, dostępne pod adresem: https://www.jpl.nasa.gov/news/news.php?feature=4541 [dostęp 29.07.2017] powrót
- [3] Harder B., Inner Earth May Hold More Water Than the Seas, National Geographic News, dostępne pod adresem: http://news.nationalgeographic.com/news/2002/03/0307_0307_waterworld.html [dostęp 29.07.2017] powrót
- [4] Ples M., Niezwykłe bariery. O błonach półprzepuszczalnych i osmozie, Biologia w Szkole, 3 (2017), Forum Media Polska Sp. z o.o., str. 52-58 powrót
- [5] Popielarska-Konieczna M., Słownik szkolny: biologia, Wydawnictwo Zielona Sowa, Kraków, 2003, str. 405 powrót
- [6] Szweykowska A., Szweykowski J., Botanika, tom 1 – Morfologia, Wydawnictwo Naukowe PWN, Warszawa, 2003 powrót
- [7] Kopcewicz J., Lewak S., Fizjologia roślin – wprowadzenie, Wydawnictwo Nukowe PWN, Warszawa, 2009, str. 44 powrót
Autorem fotografii i rysunków jest Marek Ples.
Marek Ples